
From Time Magazine to NBC Commentators: Why is everyone still so confused about lactate (and exercise physiology)?
By: Matthew L. Goodwin, PhD
September 6, 2012
The 2012 Summer Olympics have finally come and gone, and many of us were glued to our televisions, watching the world's best athletes compete. However, as has been made painfully obvious over the past weeks with reporting on the Olympic trials and the Olympics, announcers and even well-known magazines such as Time will continue to confuse viewers and readers who wish to understand the basic physiology of the world's best athletes. I cannot correct every incorrect statement here, but I hope to address a few of the big gaffes that seem to continually get reported (e.g. "lactate causes soreness") and other nonsense that has made it into widely read publications. In particular I will focus on my two areas of expertise, oxidative metabolism (1) and lactate metabolism (2), that have once again been mangled beyond recognition in recent reporting. Enjoy, and please, feel free to forward this to NBC or Time; they could use the help!
First, in the Time
magazine Summer Olympics Special issue (July 30), I read in Bill Saporito's article ("Survival of the Fittest") that "VO2max
measures milliliters of oxygen per minute per kilogram of body weight,
converted to a percentage. The higher
the number, the more oxygen you're getting to your muscles. Most of us can hit 50% on a good day. Top
college athletes clock in around 75%." There
are two significant problems with this statement. First, as
readers of this website know, VO2 simply refers to the "volume of
oxygen (O2) consumed" with VO2max being the "maximum" one
person can consume (originally verified by a plateau in oxygen consumption despite
an increase in work rate). This volume
per time can be reported however you like: in L/min or mL/min or mLkg-1min-1. Traditionally mLkg-1min-1
is used because we are usually interested in the relative ability to use oxygen (i.e. big people with more mass will
use more oxygen). So, the Time magazine writer should have left
off "converted to a percentage." That
last phrase makes the definition nonsensical.
If this were on some blog it would simply be funny, but it is Time magazine, with a weekly readership
of ~25 million. The second problem is that the writer says
that the "higher the [VO2max], the more oxygen you're getting to
your muscles"; the implication here is incorrect. VO2max is the amount of O2
consumed by the muscles, not the amount of O2 simply delivered to
the muscles. Unfortunately, the article
is full of other odd statements that are not correct, but none as egregious as
the VO2 debacle. In a more
forgivable example, Bill also reports that some events are "anaerobic" because "the distance is too short to
get oxygen pumped down from the heart." This is a bit misleading
considering that in studies that speed oxygen delivery to healthy muscle at the
onset of exercise (3), the rate at which the muscle increases its usage of
oxygen always involves a significant amount of time to ramp up to "full oxygen
usage," suggesting that something intrinsic to the muscle plays a large role in
the "adjustment" of oxygen consumption at the start of exercise. I wrote Time and corrected these mistakes, but
they neither printed the corrections nor replied to me.
But perhaps the most common "error"
in physiology reporting is the constant discussion by commentators of the
"lactic acid build up causing soreness" at sporting events. This comes up time and time again, and as
someone who has made a career of studying lactate metabolism, it is both
irritating and amazing that it has taken so long for the science to reach the
public. In fact, while my original
dissertation work focused on the control of oxidative metabolism (1), I find
myself speaking much more often about lactate metabolism. In fact, this past year I had the honor of
chairing a session and speaking at the American College of Sports Medicine Annual
meeting in San Francisco with the two lactate "gurus" of today my old mentor,
L. Bruce Gladden and George A. Brooks, from UC Berkeley. While we spoke of our recent work on lactate
and cancer, I opened with my typical "brief history" of lactate. A slightly different, adapted version is
presented below in an effort to "correct" what was said about lactate during
this Olympics. Much of this is taken almost directly from my upcoming book "The Exercise and Medicine Bible." Enjoy!
Let's start at the most basic level, with
metabolism of a muscle cell, shown below in Figure 1. Note the blood supply in red, both above and
below the circle that represents the "cell" or "muscle fiber." In yellow you see that adenosine triphosphate
(ATP) is broken down in cells to free up energy. ATP is a big molecule, so very small amounts
of it are stored; therefore, living and moving relies on continually
replenishing our ATP supply. In orange
you will see phosphocreatine (PCr), which is stored
in our cells, but also in limited amounts.
In fact, when you move rapidly, PCr breakdown
is the fastest way for your muscles to respond to energy needs. Of course even if you could use up all of the
stored ATP (you can't) and PCr, you could only fuel maximal
effort exercise for a very short time.
Typically exercise textbooks say that events that are "all out" for less
than 30 seconds rely predominantly on PCr breakdown;
this is a fair statement.
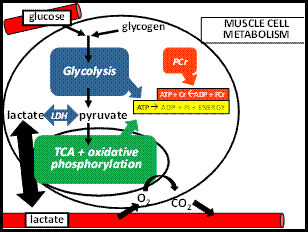
Figure
1: Schematic of muscle cell metabolism.
In blue in Figure 1 you see "glycolysis." This is a series of reactions that begins with either glucose (from the blood) or glycogen (stored within the cell). In a series of chemical reactions, more ATP is regenerated, ending invariably with lactate (more on this later). Finally, in green you see what represents mitochondria, which is where oxidative metabolism takes place (requiring oxygen). Here, pyruvate comes into the mitochondria and large amounts of ATP are regenerated, but at a cost it is a fairly slow process. In fact, in shorter events, typically the rate of ATP demand (how much energy is needed and how fast it is needed) is typically too great to rely on oxidative metabolism alone, and glycolysis that ends with lactate is preferred. In this scenario lactate is exported out into the blood (big black arrow in Figure 1) and can then circulate to be used at other places in the body.
Wait, I thought lactate was formed because of low oxygen, or "hypoxia," some might say. If you look at Figure 1 you can see that if you take oxygen away, then oxidative metabolism will not work, and indeed lactate in the blood will increase. In fact, this view of "hypoxia" causing increases in lactate is true in some circumstances (cut off blood supply to an organ) and has been taught in medical schools for years when lactate is mentioned (Figure 2). Unfortunately, this simplistic view is typically not the situation for athletes. If you were trained as an exercise science student, you likely learned about lactate metabolism in the context of Figure 3, the lactate threshold. Here you see blood lactate concentration [La] plotted vs. work rate, in this case in Watts (W) on a stationary bicycle. As the work rate increases from 30 to 60 to 90, you can see that [La] stays about the same as resting. However, as the work rate continues to rise, [La] increases exponentially. The work rate at which this happens (not the lactate concentration) is shown by the arrow in Figure 3 and is called the "lactate threshold (LT)."
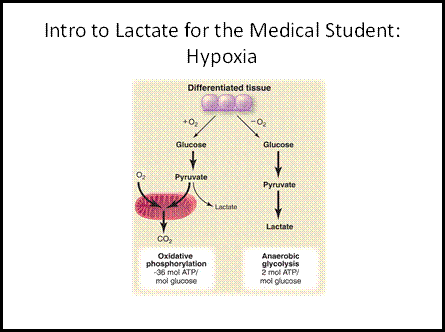
Figure
2: The traditional "medical view" of increased blood [lactate]: hypoxia (4).
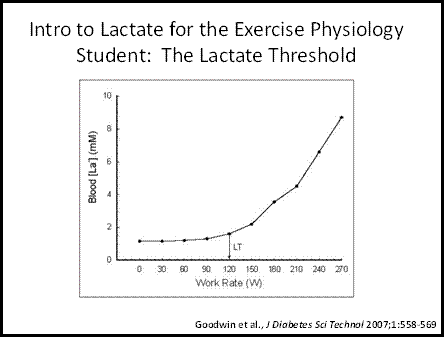
Figure
3: The typical context for learning about increased blood [lactate] for
exercise physiologists: the lactate threshold (2). Refer to text for details.
But why do we have a lactate threshold? Is it because at some work rate our hearts cannot deliver enough blood and oxygen to the muscles? Well, as pointed out earlier this is likely not the main reason, although oxygen does play a smaller (and hotly debated) role. The big reasons for having an LT are: 1) the speeded metabolism at that work rate demands "rapid" glycolysis, and thus more lactate automatically accumulates in the muscles and then in blood, 2) with higher work rates comes greater reliance on more glycolytic and fast-twitch type muscle fibers, which are built for lactate production, and 3) with higher work rates comes higher levels of catecholamines (adrenaline), which constrict vessels going to other tissues, in this case tissues like the liver that can serve to remove the accumulating lactate. Note that lactate "automatically" accumulates when glycolysis speeds up because the reaction that produces lactate from pyruvate lies heavily in the direction of lactate. Lactate concentrations are typically 10-200 times greater than the concomitant pyruvate concentrations. See below for more detail.
Speaking of the liver, you may have learned that this is the way lactate is removed from the blood. Many medical and biomedical textbooks still mention the Cori Cycle (converting lactate to glucose at the liver) as being the sole "fate of lactate." However, we now know (and we will touch more on this later) that there are many other fates of lactate. In fact, lactate serves as a very important fuel for many if not most tissues of the body, something that is now well known in exercise physiology circles and slowly spreading to the rest of medicine and the public. Figure 4 shows the "lactate shuttle," whereby lactate that is produced by some cells is then taken up and used as a fuel by both local and distant tissues. Specifically, tissues like the heart and highly oxidative skeletal muscle are excellent consumers of lactate (taking it up, converting it to pyruvate and then using the pyruvate in oxidative metabolism in the mitochondria).
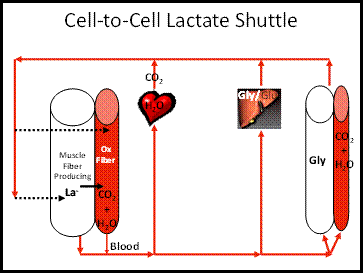
Figure
4: The lactate shuttle. On the left we see a white muscle fiber producing
lactate. This lactate then travels to
neighboring muscle tissues ("Ox Fiber") as well as distant tissues like the
heart and other muscle cells, where it is used as a fuel. In the middle you see the liver, representing
the conversion of lactate to glucose (the Cori Cycle), the traditional view
most medical students learn (Figure courtesy of LB Gladden).
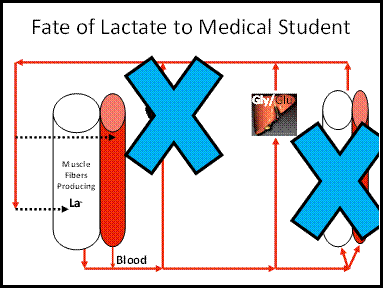
Figure
5: Instead of the lactate shuttle, most medical students learn the incorrect
view that conversion of lactate to glucose at the liver (the Cori Cycle) is the
only "fate" of lactate (Figure courtesy of LB Gladden).
As you may be aware, particularly from listening to commentators on NBC, lactate has for a long time been getting a "bad rap." In fact, when George Brooks was interviewed by The New York Times in 2006 (5), he called this idea that lactate is bad "one of the classic mistakes in the history of science. I tend to agree. A quick look back over history gives us the reasons lactate was viewed in this way. Hang in there through this short history lesson - it'll come together and make sense in the end! First, lactic acid was discovered by Carl Wilhelm Scheele in 1780 when he isolated it from soured milk (and hence the name). But it wasn't until 1807-08 that we began to associate elevated lactate levels with fatiguing exercise (when Jöns Jacob Berzelius reported lactate in the muscles of hunted stags) (6). This idea that lactate might be "bad" and "fatigue-causing" only gained more traction when the great scientist Otto Meyerhof (who actually isolated the enzyme(LDH) that converts pyruvate to lactate or vice versa) did experiments on isolated frog legs. In these experiments he contracted the muscles until they fatigued, and then he examined the fluid the muscles were bathed in. When he did this he found that lactate in the solution bathing the muscles was very high! Finally, to complete the early picture of lactate being "bad" and a "hypoxic byproduct," we must mention that in 1857 Louis Pasteur showed us that yeast broth fermented when oxygen was low, thus beginning our understanding of fermentation as a direct result of low oxygen (in his case it was pyruvate to ethanol; in human muscle the equivalent is pyruvate to lactate, shown in Figure 1). Gladden has called the period beginning with Meyerhof (1930s-1970s) the "dead-end waste product era" of lactate (7), as many viewed it as a waste product. To this day it is hard for many to understand that low oxygen is often the exception rather than the rule in day-to-day production of lactate in our bodies.
While many studies hinted that lactate was not all bad before 1968, it was then that Wendell Stainsby, a physiologist at the University of Florida, performed a series of studies that would change our fundamental understanding of lactate metabolism. In a 1966 paper (8), Stainsby noted that when he used his isolated canine muscle to study contractions, there was a transient lactate output from the muscle for the first 5-20 min of contractions. He then set out to discover why this was happening, at first investigating whether the muscle was hypoxic at the onset of contractions. In 1968 he published the landmark paper (9) that showed that the contracting, lactate-producing canine muscle was not dysoxic! (Dysoxia = O2 low enough to impair oxidative metaboplism.) At the time this paper was argued about a great deal, and some specifics of the methods have been hotly debated, but it opened the flood gates for experiments examining lactate and our current understanding of it as a valuable fuel, not a deadend waste product. In fact, if you examine the lactate-dehydrogenase (LDH) reaction (lactate ßàpyruvate) specifically, you will see that it is a "near-equilibrium" reaction this means that any time there is pyruvate there is lactate it reaches equilibrium. To that end, that "equilibrium" lies heavily toward lactate, meaning a tissue might be "at equilibrium" with 1 pyruvate for 100 lactate. In other words, regardless of oxygen status, lactate is being formed.
Cooling Down to Flush Lactic Acid?
At this point you might think "So lactate accumulates when oxygen is plentiful, like during strenuous exercise; so what? Either way, all I hear about is how I must cool down after I exercise to get the lactic acid out. Otherwise I will be really sore." Every time I think this myth is finally dead, some coach, athlete, writer, or sports commentator demonstrates to me that it is alive and well. Figure 6 shows the typical [La] in the blood following exercise that generates the highest levels of lactate (30-90 seconds of all out exercise). The time shown is in minutes post-exercise; when the 30-90 second bout is finished, lactate in the arterial blood peaks 3-8 min later, with some reporting levels as high as 25mM (shown with "X" in Figure 6). However, note that within 1 hour, blood [La] is almost completely back to resting levels! Yes, if you perform an active, low intensity cool down the slope of this line changes slightly, but whether you are active or just sit, [La] will certainly be back to normal within 2 hours. In contrast, most soreness after exercise is felt 24-48 hours after a bout of exercise, when [La] has been back to normal for some time! Further, the types of exercises that induce the most soreness typically do not alter lactate levels very dramatically (eccentric weight lifting in which the muscle lengthens while it contracts). So next time someone tells you to cool down to get the lactic acid out, point out that 1) lactate levels in the blood will be back to resting within ~ an hour regardless, 2) it doesn't cause soreness, and 3) exercise that causes soreness typically doesn't cause very high [La] (and vice versa).
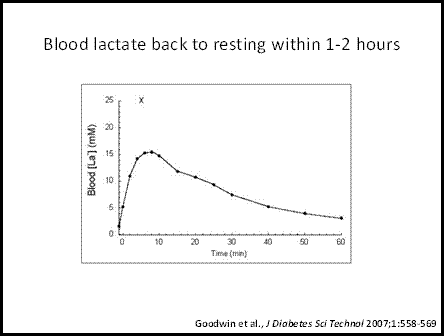
Figure
6: Representative example of post-exercise blood [lactate]. Note that this is blood [lactate] immediately
after 30-90 seconds of "all-out," maximal exercise (the exercise that generates
the highest blood lactate concentration.
Importantly, note that lactate levels are almost completely back to resting
within 1 hour (and are totally back to normal within 2 hours) (2).
So is lactate good or bad? It doesn't cause soreness and it seems to be produced when there is plenty of oxygen; so what is the point? Well, in the late 1960s Depocas and colleagues performed a series of studies with "labeled" lactate molecules that were infused into humans at rest and during exercise (10). Despite the traditional view that lactate must "go to the liver to be converted to glucose via the Cori Cycle," they showed that ~50% of the lactate infused at rest and ~75% infused during exercise was directly oxidized as a fuel. The interpretation is that conversion of lactate to glucose in the liver was not the primary fate of lactate. Instead, most of the lactate was going to other tissues where it was being converted to pyruvate and metabolized as a fuel in the mitochondria. George Brooks and colleagues followed up on this in the 1970s with studies examining rats run to exhaustion with the same result most of the lactate was again used as a fuel (oxidized)(11)! In fact, Brooks continued his investigations (and continues today) and in 1984 proposed the "lactate shuttle hypothesis," which is now well known throughout muscle physiology and making inroads into tumor physiology (12, 13). The lactate shuttle hypothesis has now been so clearly supported by the tremendous amount of data that many now call it "a theory."
Briefly, the lactate shuttle espouses that lactate is the major way whole body metabolism seems to be coordinated, with some cells producing lactate and releasing it into the blood while other cells take it up and use it as a fuel. In fact, during moderate exercise, lactate flux can exceed glucose flux, further underscoring its importance as a fuel. We now know that lactate is a valuable fuel for any tissue with mitochondria (i.e. almost all cells in humans besides red blood cells, a few cells in the eye, etc.), serving as a valuable fuel for even the heart and brain! Figure 7 is from one of my favorite articles, a Science paper from 1988 (14). In this experiment, Avital Schurr and colleagues examined brain tissue under two different conditions (open circles and closed circles). In both conditions they first removed the glucose supply, after which both tissues began to fail, as evidenced by "spike amplitude" going to 0 (Figure 7). Then, they raised lactate to 10mM in one group, but not the other. The lactate-supplied tissue made a full recovery, providing evidence that lactate is such a good fuel that perhaps it can even replace glucose. This latter statement has been the basis for some of my ongoing collaborative work, in which our ultimate goal is to starve tumors by removing glucose at the same time we provide lactate to fuel healthy tissues (most tumor cells heavily prefer glucose over lactate), something we have been calling "lactate-protected hypoglycemia (LPH)" (well beyond the scope of this review).
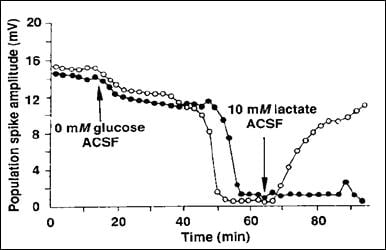
Figure
7: Lactate replacing glucose as a fuel in mammalian hippocampus slices. Two groups of tissues were studied in vitro. In both groups glucose was removed from
solution (shown with arrow and "0 mM glucose"). Both tissues began to fail (spike amplitude
falling to 0). In one group nothing was
changed after that (closed circles); in the other group (open circles) 10 mM lactate was added.
The lactate fully replaced glucose as a fuel and the tissue returned to
normal vitality (14).
To conclude, I hope you've found this both informative and interesting. You will notice that I left out the important question "does lactate cause fatigue?" The main reason is that while I would argue that much of the recent data suggest "no," there has been no final resolution. For example, when you form lactate you are contributing a strong ion that serves to lower the pH of solution, and low pH has been implicated in fatigue. (Formation of lactate contributes to a lower pH, regardless of what some might tell you about "consuming a proton in the LDH reaction." You need to look at the larger solution you are examining, not the isolated reaction, to meaningfully comment on the acid-base status. For example, when patients metabolize lactate from Ringers' lactate (a common resuscitation solution), blood pH increases as the lactate is metabolized; this has been demonstrated many different ways experimentally. However, there are a few physiologists out there who continue to argue that forming lactate "prevents" acidosis - I think the evidence is against them.) Regardless, there is some reasonable evidence that protons (what has increased in solution to make a solution have a "low pH") contribute to the fatigue process, and perhaps even the "burning" one feels during hard exercise. However, when many of the studies that showed that protons "caused fatigue" were corrected to physiological temperatures (~30°C instead of ~0°C), there was little to no effect on contractions, suggesting little role in fatigue. (Be warned: not all studies show this and some very good physiologists will argue that protons are indeed often the cause of fatigue.) Regardless of the role of protons in fatigue, lactate itself likely plays only a very small role, as numerous studies have been done infusing lactate into animals with little apparent effect on contractions. However, as I tell the first year medical students each year at Cornell, fatigue is still not well understood, and studies are ongoing to elucidate exactly what causes fatigue. Lactate would certainly not be at the top of my list.
All in all, I hope you are able to take away a few salient points from this article. To quickly recap: 1) Maximum oxygen consumption is a number (usually corrected for body weight), not a percentage; 2) lactate is a valuable fuel, not a waste product; 3) lactate concentration increasing in the blood during intense exercise is typically not due to lack of oxygen, but rather speeded glycolysis, increased reliance on glycolytic muscle fibers, and decreased blood flow to lactate removal sites; 4) lactate does not cause soreness; and 5) lactate levels in the blood are back to resting within 2 hours regardless of what you do post-exercise. And remember, don't believe everything you hear on TV or read in magazines particularly if it has to do with exercise physiology or lactate metabolism. After reading this it's likely that you know more than they do!
References:
1. Goodwin ML et al. VO2 on-kinetics in isolated canine muscle in situ during slowed convective O2 delivery. J Appl Physiol 112(1); 9-19, 2012.
2. Goodwin ML et al. Blood lactate measurement and analysis during exercise: a guide for clinicians. J Diabetes Sci Technol 1(4); 558-69, 2007.
3. Grassi B et al. Faster adjustment of O2 delivery does not affect VO2 on-kinetics in isolated in situ canine muscle. J Appl Physiol 85(4); 1394-403.
4. Vander Heiden MG et al. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science 324(5930); 1029-33.
5. Kolata G. Lactic acid is not muscles' foe, it's fuel. The New York Times, May 16, 2006.
6. Gladden LB. 200th anniversary of lactate research in muscle. Exer Sport Sci Rev 36(3); 109-15, 2008.
7. Gladden LB. Lactate metabolism: a new paradigm for the third millennium. J Physiol 558(Pt 1); 5-30, 2004.
8. Stainsby WN & Welch HG. Lactate metabolism of contracting dog skeletal muscle in situ. Am J Appl Physiol 211; 177-83, 1966.
9. Jobsis FF & Stainsby WN. Oxidation of NADH during contractions of circulated mammalian skeletal muscle. Resp Physiol 4; 292-300, 1968.
10. Depocas F et al. Rates of formation and oxidation of lactic acid in dogs at rest and during moderate exercise. Can J Physiol Pharmacol 47(7); 603-10, 1969.
11. Brooks GA et al. Glycogen synthesis and metabolism of lactic acid after exercise. Am J Physiol 224(5); 1162-6, 1973.
12. Semenza GL. Tumor metabolism: cancer cells give and take lactate. J Clin Invest 118(12); 3835-7, 2008.
13. Gladden LB et al. Fuel for cancer cells? Cell Cycle 10(15); 2421-2, 2011.
14.
Schurr A et al.
Lactate-supported synaptic function in the rat hippocampal slice preparation. Science 240(4857); 1326-8, 1988.
Matthew L. Goodwin was a D1 runner in college and completed his PhD in lactate expert L. Bruce Gladden's lab, where he studied lactate metabolism and oxidative metabolism. He completes his MD from Cornell next spring, after which he will be pursuing Orthopaedic Surgery as a career. He currently teaches about muscle physiology and physical activity at Weill Cornell Medical College and recently published the textbook "Biomedical Physiology." His newest book, "The Exercise and Medicine Bible," aims to clarify confusion about topics in exercise and medicine; it's due out in 2013. He can be reached atMatthewLGoodwin@gmail.com and on twitter @Physiologyguy.
Comments, questions, suggestions, or a story you'd like to submit? Email us.
![]()
|
|
|
|
|
|